



Specifičnost detekcije
U većini slučajeva, svrha dizajna prajmera je maksimiziranje specifičnosti PCR-a.Ovo je određeno manje-više predvidljivim uticajem mnogih varijabli.Jedna važna varijabla je sekvenca na 3′kraju prajmera.
Važno je da je veća vjerovatnoća da će PCR testovi dizajnirani za specifičnost održati visoku efikasnost u širokom dinamičkom rasponu, jer test ne proizvodi nespecifične produkte amplifikacije, čime se nadmeće sa PCR reagensima ili inhibira glavnu reakciju amplifikacije.
Naravno, u nekim slučajevima specifičnost nije najvažnija, na primjer, kada je cilj kvantificiranje blisko povezanih, ali različitih patogena, potrebni su posebni standardi dizajna, optimizacije i verifikacije.
Kriva topljenja je standardna metoda za procjenu specifičnosti amplikona, barem u smislu da li treba pojačati jednu metu.Međutim, mora se naglasiti da krivulje topljenja mogu biti pogrešne jer, na primjer, na njih mogu utjecati kombinirani efekti suboptimalnih prajmera i niske koncentracije šablona.
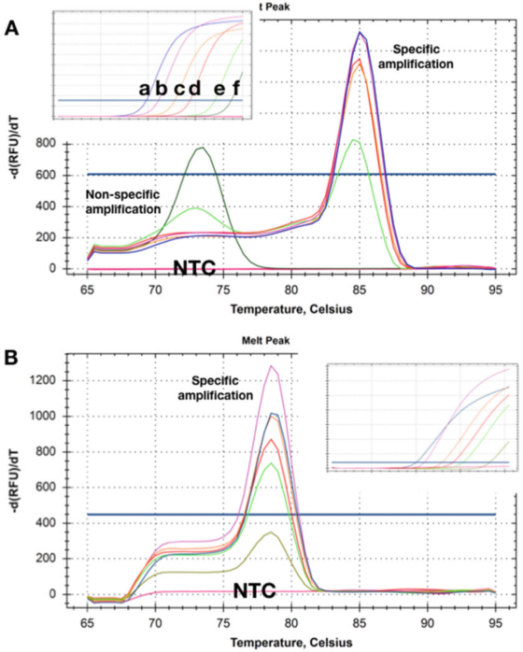
P5 |Kriva topljenja pokazuje Tm pomake dobijene iz dvije detekcije različitih količina dvije ciljne DNK.
A. Pri višim koncentracijama (ad)), nema očiglednog dimera prajmera nakon što je qPCR merenje završeno.Kako se koncentracija šablona smanjuje na 50 kopija (e), počinje se pojavljivati nespecifičan proizvod i postaje jedini proizvod s najnižom koncentracijom (f).
B. Test je zabilježio isti Tms na svim ciljnim koncentracijama, a nije bilo očiglednog dimera prajmera čak ni pri najnižoj koncentraciji (5 kopija).Kada se koriste ove dvije metode detekcije, u NTC-ovima nisu otkriveni proizvodi pojačanja.
P5 prikazuje krivulje rastvaranja dobivene s uzorcima u kojima je šablon prisutan u različitim koncentracijama.P 5a pokazuje da su pri dvije najniže koncentracije Tms proizvedenih nespecifičnih produkata amplifikacije niži od onih specifičnih amplikona.
Očigledno, ova metoda detekcije ne može se pouzdano koristiti za otkrivanje ciljeva koji postoje u niskim koncentracijama.
Zanimljivo je da NTC, tj. uzorci bez DNK uopšte, nisu zabeležili (nespecifične) proizvode amplifikacije, što ukazuje da pozadinska genomska DNK može učestvovati u nespecifičnoj amplifikaciji/polimerizaciji.
Ponekad se takvi osnovni prajmeri i nespecifična amplifikacija ne mogu popraviti, ali je često moguće dizajnirati metod detekcije koji nema nespecifično pojačanje u bilo kojoj koncentraciji šablona i NTC (P 5b).
Ovdje će čak i snimanje pojačanja ciljne koncentracije sa Cq od 35 proizvesti specifičnu krivulju rastvaranja.Slično, NTC nisu pokazivali znakove nespecifične amplifikacije.Ponekad, ponašanje detekcije može zavisiti od matične tečnosti, a samo nespecifična amplifikacija se detektuje u određenim puferskim sastavima, što može biti povezano sa različitim koncentracijama Mg2+.
Stabilnost detekcije
Optimizacija Ta je koristan korak u empirijskoj verifikaciji i procesu optimizacije qPCR detekcije.Pruža direktnu indikaciju robusnosti seta prajmera tako što pokazuje temperaturu (ili temperaturni raspon) koja proizvodi najniži Cq bez pojačavanja NTC-a.
Dvostruka do četverostruka razlika u osjetljivosti možda nije važna za ljude s visokom ekspresijom mRNA, ali za dijagnostičke testove može značiti razliku između pozitivnih i lažno negativnih rezultata.
Ta svojstva qPCR prajmera mogu se značajno razlikovati.Neki testovi nisu jako robusni i ako se ne izvode pod optimalnom Ta vrijednošću prajmera, brzo će se srušiti.
Ovo je važno jer je ova vrsta detekcije često problematična u stvarnom svijetu, a čistoća uzorka, koncentracija DNK ili prisutnost druge DNK možda nisu optimalni.
Osim toga, ciljni broj kopija može varirati u širokom rasponu, a reagensi, plastični pribor ili instrumenti mogu se razlikovati od onih koji se koriste prilikom postavljanja testa.
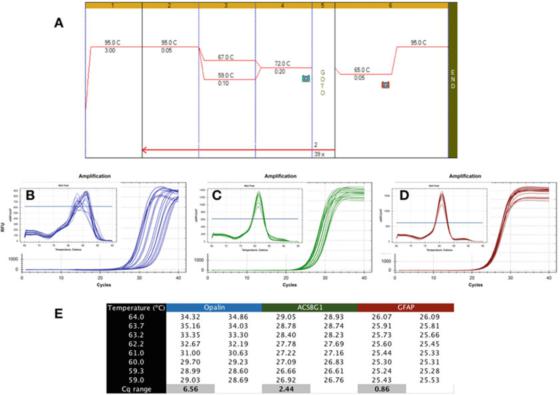
P6|Gradijent temperature pokazuje različitu robusnost PCR detekcije.
A. Koristite Biolineov Sensifast SYBR mastermix (kataloški broj BIO-98050) za izvođenje PCR-a na cDNK pripremljenoj od RNK ljudskog mozga.
B. Koristite Bio-Rad CFX qPCR instrument da snimite mapu amplifikacije i krivulju rastvaranja apalena (NM_033207, F: GCCATGGAGGAAAGTGACAGACC, R: CTCATGTGTGGGTGATCTCCTAGG).
C. Grafikon pojačanja i kriva topljenja ACSBG1 (NM_015162.4, F: CTACACTTCCGGCACCACTGG, R: GTCCACGTGATATTGTCTTGACTCAG).
D. Grafikon amplifikacije i kriva rastvaranja GFAP-a (NM_002055.5, F: TGGAGAGGAAGATTGAGTCGCTGG, R: CGAACCTCCTCCTCGTGGATCTTC).
E. Cqs snimljeni na različitim temperaturama žarenja, pokazujući razliku u Cq zabilježenoj pod temperaturnim gradijentom od 7C.
P 6 pokazuje tipičan rezultat nepoželjnog testa, gdje je qPCR izveden korištenjem gradijenta Tas između 59C i 67C (P 6a), koristeći prajmere za tri gena specifična za ljudski mozak.
Iz grafikona amplifikacije se može vidjeti da su opalin prajmeri daleko od idealnih jer je njihov optimalni Ta raspon vrlo uzak (Slika 6b), odnosno, Cqs su široko raspršeni, što rezultira značajnom usporedbom Cqs sa svojim optimalnim Cqs Low.
Ova metoda detekcije je nestabilna i može dovesti do suboptimalnog pojačanja.Stoga bi ovaj par prajmera trebalo redizajnirati.Osim toga, analiza krivulje topljenja (umetnuti) pokazuje da specifičnost ove metode detekcije također može biti problematična, jer je kriva topljenja svakog Ta različita.
Metoda detekcije ACSBG1 prikazana u P 6c je robustnija od metode detekcije Opalina iznad, ali je još uvijek daleko od idealne i vjerovatno je da se može poboljšati.
Međutim, naglašavamo da ne postoji neophodna veza između robusnosti i specifičnosti, jer krivulja rastvaranja proizvedena ovom metodom detekcije pokazuje istu vršnu vrijednost u svim Tas (umetnutim).
S druge strane, test robusnosti je mnogo tolerantniji, proizvodeći slične Cqs u širokom rasponu Tas, kao u GFAP testu prikazanom u P 6d.
Razlika u Cqs dobijenoj u istom opsegu od 8 stepeni Celzijusa je manja od 1, a kriva rastvaranja (umetnuti) potvrđuje karakteristike detekcije u ovom temperaturnom opsegu.Vrijedi napomenuti da izračunati Tas i stvarni Ta raspon mogu biti vrlo različiti.
Postoje mnoge smjernice osmišljene da pomognu istraživačima da dizajniraju efikasne prajmere, od kojih se većina zasniva na davno utvrđenim pravilima i puno pažnje je posvećeno 3′kraju prajmera.Često se preporučuje uključivanje G ili C na kraju od 3' i dvije G ili C baze (GC stezaljka), ali ne više od dvije od posljednjih 5 baza.
U praksi, ova pravila mogu voditi istraživače, ali nisu nužno tačna u svim okolnostima.
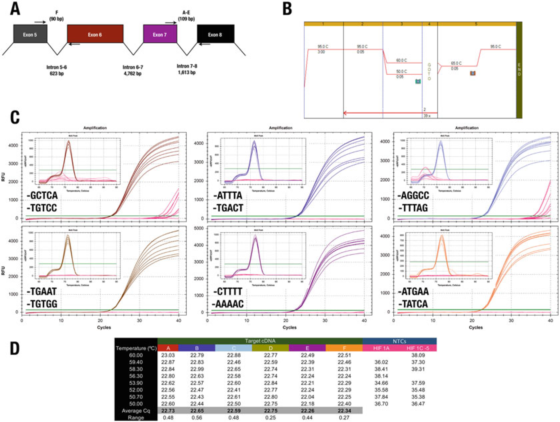
P7 |3′kraj prajmera ima mali uticaj na specifičnost ili efikasnost.
A. Položaj prajmera za ljudski HIF-1α (NM_181054.2) gen.
B. Koristite Agilent Brilliant III SYBR Green matičnu tečnost (kat. br. 600882) da pojačate šest testnih jedinica.
C. Grafikon amplifikacije i krivulja topljenja snimljeni Bio-Rad-ovim CFX qPCR instrumentom i 3′end prajmerima.NTC su prikazani crvenom bojom.
D. Cqs zapis svake ispitne stavke
Na primjer, rezultat u P 7 je u suprotnosti s pravilom 3′kraja.Svi dizajni daju u osnovi iste rezultate, sa samo dvije kombinacije prajmera koje dovode do nespecifičnog pojačanja u NTC.
Međutim, ne možemo podržati učinak GC klipa, jer u ovom slučaju korištenje A ili T kao maksimalno 30 baza ne smanjuje specifičnost.
Test C, gdje F prajmer završava na GGCC, je zabilježio Cqs u NTC, što ukazuje da bi se možda željele izbjeći ove sekvence na 30-kraju.Naglašavamo da je jedini način da se odredi najbolja 3′krajna sekvenca para prajmera da se eksperimentalno procijene neki kandidati za prajmere.
Efikasnost pojačanja
Važno je da iako nespecifična PCR detekcija nikada ne može postati specifična, efikasnost amplifikacije se može prilagoditi i maksimizirati na mnogo različitih načina promjenom enzima, matične tekućine, aditiva i uvjeta ciklusa.
Za procjenu efikasnosti PCR detekcije, najbolje je koristiti serijsko razrjeđenje od 10 ili 5 puta veće od ciljne nukleinske kiseline, odnosno „metoda standardne krive“.
Ako se PCR amplikoni ili sintetičke DNK mete koriste za generiranje standardne krive, serijska razrjeđenja ovih meta treba pomiješati sa konstantnom količinom pozadinske DNK (kao što je genomska DNK).
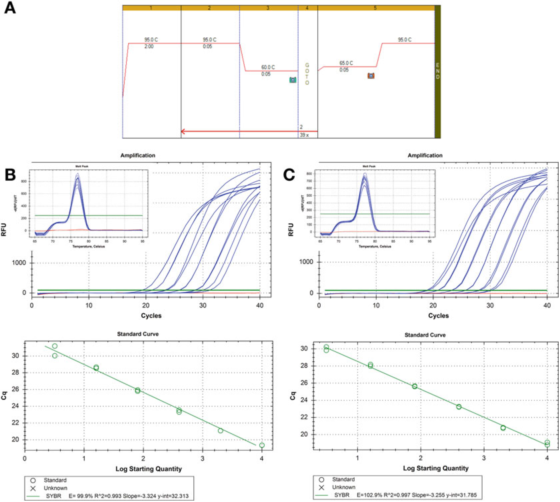
P8 |Kriva razblaženja za procenu efikasnosti PCR.
A. Koristite prajmere za HIF-1: F: AAGAACTTTTAGGCCGCTCA i R: TGTCCTGTGGTGACTTGTCC i Agilentov Brilliant III SYBR Green mastermix (kataloški broj 600882) za PCR i uslove krive topljenja.
B. 100 ng RNK je reverzno transkribovano, razrijeđeno 2 puta, a serijski razrijeđeni uzorci cDNK razrijeđeni su 5 puta do 1 ng ljudske genomske DNK.Kriva topljenja je prikazana na umetku.
C. RT reakcija, razblaživanje i serijsko razblaživanje su ponovljeni za drugi uzorak cDNK, a rezultati su bili slični.
P 8 prikazuje dvije standardne krive, koristeći istu metodu detekcije na dva različita uzorka cDNK, rezultat je ista efikasnost, oko 100%, a slična je i vrijednost R2, odnosno stepen uklapanja eksperimentalnih podataka i linije regresije ili stepena linearnosti podataka.
Dvije standardne krive su uporedive, ali nisu potpuno iste.Ako je svrha precizno kvantificirati cilj, mora se napomenuti da je neprihvatljivo dati izračun broja kopija bez objašnjenja nesigurnosti
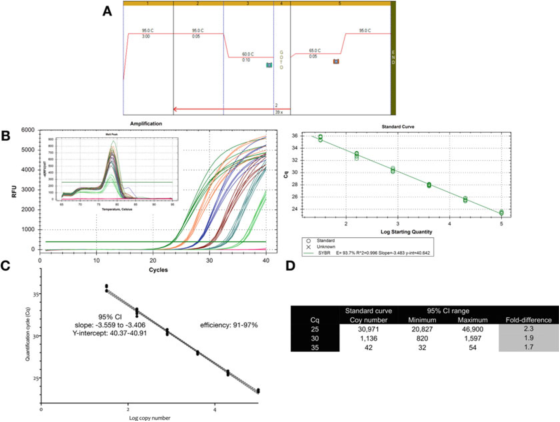
P9 |Mjerna nesigurnost povezana s kvantifikacijom koristeći standardnu krivu.
A. Koristite prajmere za GAPDH (NM_002046) da izvršite PCR i uslove krive topljenja.F: ACAGTTGCCATGTAGACC i R: TAACTGGTTGAGCACAGG i Biolineov Sensifast SYBR mastermix (kataloški broj BIO-98050).
B. Tabela pojačanja, kriva topljenja i standardna kriva snimljena Bio-Rad-ovim CFX qPCR instrumentom.
C. Grafikon standardne krive i 95% interval pouzdanosti (CI).
D. Broj kopija i interval pouzdanosti od 95% tri vrijednosti Cq izvedene iz krive razrjeđenja.
P 9 pokazuje da je za optimizirani test inherentna varijabilnost jedne standardne krive približno 2 puta (95% interval pouzdanosti, minimum do maksimuma), što može biti najmanja varijabilnost koja se može očekivati.
Povezani proizvod:
Animal Tissue Direct PCR komplet
Vrijeme objave: Sep-30-2021



